Source: http://www.tulane.edu/~wiser/protozoology/notes/vector.html
Марк Ф. Визер
Паразитите кои престојуваат во крвта или внатрешните органи на домаќинот имаат логистички проблеми во однос на инфицирање на нов домаќин. За разлика од фекално-оралното пренесување, каде што инфективните фази се излачуваат во околината, потенцијалните нови домаќини не би имале нормален контакт со паразитот. (Во еволутивна смисла, преносот со трансфузија на крв би бил неодамнешен настан.) Преносот предатор-плен е една стратегија што ја користат протозоите, како што се Токсоплазма и Саркоцистис, за да ги надминат овие пречки за пренос. Како што имплицира името, преносот на плен предатор вклучува два различни домаќини. Предаторот ја стекнува инфекцијата јадејќи заразен плен. Ова ќе предизвика цревна инфекција кај предаторот и ќе резултира со екскреција на инфективни фази со измет. Пленот, генерално тревојади, ќе се зарази јадејќи ги инфективните фази што ги среќава во околината. После ингестија од соодветен плен, паразитот ќе го премине цревниот епител и ќе зарази внатрешни органи или ткива во рамките на домаќинот, каде што чека следниот предатор да го проголта својот плен. [Содржина на страницата]
Векторското пренесување е друга стратегија што ја користат протозоините паразити кои живеат во крвта или внатрешните ткива во рамките на неговиот домаќин. Оваа стратегија вклучува хематофог (т.е. хранење со крв) членконога што служи како посредник помеѓу последователни домаќини на ‘рбетници. Неколку човечки болести предизвикани од протозои се пренесуваат со различни вектори на членконоги (Табела). Векторите не се едноставно „летечки шприцеви“, туку претставуваат втор домаќин на протозоаниот паразит. Така, животниот циклус на пренесени векторски болести вклучува и комплексни интеракции протозоа-векторска интеракција аналогно на сложените интеракции меѓу протозоите и луѓето. Векторското пренесување веројатно еволуирало повеќе пати.
| Пренесена протозоа од вектор | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||||||||||||||||||
Векторското пренесување вклучува и комплексни интеракции помеѓу луѓето и векторите. Ова вклучува биологија на интеракции меѓу човечки членконоги, како и еколошки размислувања. Така, параметрите пренесени од вектори покажуваат сложени животни циклуси кои вклучуваат интеракции помеѓу луѓето, протозоите и членконогите. Биологијата на вектори и нивните интеракции со луѓето обезбедуваат можни средства за контрола на преносот на овие болести.
- Це-це и африкански трипанозоми
- Тријатомини и Трипаносома крузи
- Песочни мушички и Леишманија
- Анофелес и Плазмодиум
- Крлежи и Бабезија
- Стратегии за контрола
Це-це и африкански трипаносоми
 Родот Глосина, вообичаено име Це-це, содржи најмалку 30 видови или подвидови муви кои се наоѓаат во субсахарска Африка. Возрасните Це-це се жолти, кафеави или црни муви долги 6-14 мм. Дијагностички карактер на родот е ‘ќелијата клетка’ пронајдена во центарот на крилјата (Слика). Повеќето видови спаѓаат во една од двете главни групи претставени од Г. палпалис и Г.
Родот Глосина, вообичаено име Це-це, содржи најмалку 30 видови или подвидови муви кои се наоѓаат во субсахарска Африка. Возрасните Це-це се жолти, кафеави или црни муви долги 6-14 мм. Дијагностички карактер на родот е ‘ќелијата клетка’ пронајдена во центарот на крилјата (Слика). Повеќето видови спаѓаат во една од двете главни групи претставени од Г. палпалис и Г. 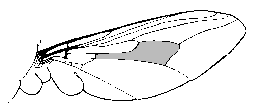 морситани. Групата палпалис е поврзана со речни екологии и често се наоѓа во близина на потоци, реки и езера во западна и централна Африка. Групата Морситани најчесто се поврзува со шуми на савана и земја со сува грмушка во источна Африка. Групите палпалис и морситани се поврзани со преносот на Трипаносом гамбиенс и T. родесиенсе, соодветно. Разликите во екологијата и интеракцијата со резервоарите на овие два вида на Це-це придонесуваат за различните манифестации на болести предизвикани од двата африкански видови трипанозми (Видете ја табелата во белешките за африканските трипанозоми.)
морситани. Групата палпалис е поврзана со речни екологии и често се наоѓа во близина на потоци, реки и езера во западна и централна Африка. Групата Морситани најчесто се поврзува со шуми на савана и земја со сува грмушка во источна Африка. Групите палпалис и морситани се поврзани со преносот на Трипаносом гамбиенс и T. родесиенсе, соодветно. Разликите во екологијата и интеракцијата со резервоарите на овие два вида на Це-це придонесуваат за различните манифестации на болести предизвикани од двата африкански видови трипанозми (Видете ја табелата во белешките за африканските трипанозоми.)
И машкото и женското Це-це се незаситни хранилки и важните видови вектори имаат тенденција да се хранат со големи цицачи. Це-це значи „мува деструктивна за добитокот“ во Сехуана (оттука и Це-це „мува“ е излишна). Каснувањето од Це-це е болно, но обично има помали последици. Повремено ќе се формира заварување, а некои лица ќе станат чувствителни на плунката. Сепак, Це-цевите се постојани колибри и можат да бидат доста отежнувачки за луѓето и животните. Це-цевите се „базени“ колибри (т.е. телмофаги) кои ја чистат кожата со делови од устата, а потоа ги проголтуваат крвта и лимфата што се влеваат во површната лезија. Преносот на трипанозом се случува преку плунка која се инјектира во местото на каснување. Плунката содржи супстанции кои ги шират крвните садови и спречуваат коагулација.
На животниот циклус на африкански трипаносоми вклучува наизменични помеѓу ‘рбетник домаќин и на Це-це вектор. Трипомастиготите во крвотокот на ‘рбетниот домаќин се проголтани од Це-це и се претвораат во проциклични трипомастиготи кои се реплицираат во цревата на Це-цетот. За да се заврши животниот циклус, овие проциклични форми мора да се пробијат до плунковните жлезди. Не е познат точниот механизам со кој паразитот мигрира од Це-цевиот црев во плунковните жлезди. Предложени се два начина: 1) класична патека по која паразитот „се повлекува“ низ дигестивниот систем и мигрира кон плунковниот канал или 2) директната патека по која паразитот продира во перитрофичната мембрана и епителот на цревата за да добие пристап до хемолимфа. Било кој пат претставува препреки и ги илустрира сложените интеракции помеѓу протозоаниот паразит и векторот (Слика).
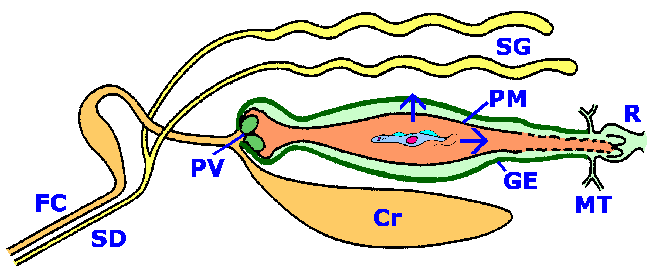
Шематски приказ на интеракции на трипанозом-Це-це. Трипомазиготите се земаат заедно со крвниот оброк и минуваат низ каналот за храна (FC) и посевите (Cr). Крвниот оброк е обвиткан во перитрофичната мембрана (РМ) од провентрикуларниот вентил (PV) кога влегува во средното црево. Миграцијата на процикличните трипомастиготи од средното црево кон плунковните жлезди (SG) вклучува два можни правци (прикажани со стрели): „класичниот“ и „краток спој“. Класичната патека предизвикува миграција кон задниот дел од цревата (малпигиски цевки, МТ и ректум, R) и излегување од перитрофичната мембрана. Процикликот потоа мигрира напред преку ектоперитрофичниот простор, излегува на провентрикуларниот вентил, поминува низ каналот за храна и влегува во плунковниот канал (SD) за да добие пристап до плунковните жлезди. Патеката на краток спој вклучува директно продирање на перитрофичната мембрана и епител на цревата (GE) за да се добие пристап до хемоцелот. Од хемоцелот, трипанозомите ги наоѓаат и продираат во плунковните жлезди.
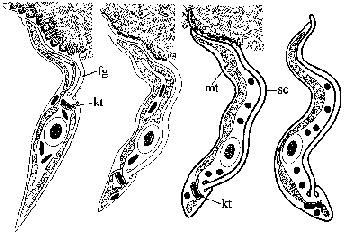
Паразитот се претвора во форма на епимастиготи откако ќе стигне до плунковната жлезда. Епимазигот се прицврстува на епителните клетки на плунковните жлезди преку својот флагелум (fg) и се подложува на понатамошно реплицирање. Диференцијацијата во метациклични трипомастиготи вклучува појава на површински слој (sc) и промени во митохондриите (mt) за да се придружува миграцијата на кинетопластот (kt) до задниот крај (Слика). Овие развојни промени се неопходни подготовки за да станете инфективни кај домаќинот на цицачите. Зрелиот метацикличен трипомастигот се одвојува од епителните клетки и останува во луменот на плунковната жлезда сè додека не се исфрли за време на хранењето со Це-це.
(Вратете се во Животниот циклус на Африканскиот трипанозом или одете на Контролни стратегии.)
Тријатомини и Трипаносома крузи

Слика изменета од А.М. Роуз (http://www.arose.net/triatoma/pics.htm) и се користи со дозвола (© H Knüttel & AM Rose).
Неколку родови и видови на триатомини се способни да пренесат Трипаносома крузи. Најважните вектори се: Триатома инфестанци, Панстронгилус мегистас и Родниус проликс. Тријатомините се генерално големи инсекти со големина од 5-30 мм. Тие се првенствено инсекти од Новиот свет и се протегаат од Аргентина преку САД. Theивотниот циклус се состои од пет нимфални години, проследени со сексуално зрели возрасни лица. Само возрасните имаат крилја. Тријатомините се хранат со крв низ целиот живот и сите фази можат да се заразат со T. крузи. Заедничките имиња вклучуваат бубачка-атентатор, грешка за бакнување (во врска со неговата тенденција да гризат околу лицето) и бубачка со коноза (во однос на нивните зашилени глави).
Повеќето триатомини не се специфични за домаќинот и се хранат со широк спектар на цицачи, како и птици и влекачи. Пренесувањето на T. крузи често се поврзува со домородни видови кои се прилагодиле на живеење во човечки живеалишта. Таквите живеалишта се генерално колиби од кирпич или покриви од слама, кои обезбедуваат многу скривалишта за грешките. Грешките излегуваат навечер за да се хранат со луѓе што спијат. Тие се капиларни колибри и времето на хранење се движи од 3-30 минути. Возрасните можат да достигнат до 0,25 мл крв за време на хранење, а вкупно 4-10 мл крв од ‘рбетници може да се потрошат од грешка на триатомин за време на нејзиниот живот. Каснувањата обично се безболни, и покрај големиот пробосцис. Се претпоставува дека плунката содржи анестезија. Иако хранењето со триатомин предизвикува мала болка, нивните каснувања можат да предизвикаат непријатна лезија на местото на хранење и може да предизвикаат реакција на преосетливост.
Тријатомините се инфицираат кога трипомастиготите во крвта се проголтаат со крвниот оброк. Трипомазиготите се претвораат во епимастиготи кои се репликативна форма која се наоѓа првенствено во средното црево. Епимастиготите ќе се развијат во неразделни метациклични трипомазиготи. Метациклогенезата е скоро ексклузивна за ректумот и е во корелација со приврзаноста кон ректалниот епител. Интеракциите вектор-паразит се важни за успешна репликација и диференцијација на T. крузи. (Видете исто така: Колиен и Шауб, Развој на Трипаносома крузи во триатоминеа. Паразитол. Денес 16:381; 2000 г.) Инфективните метациклични трипомастиготи се излачуваат со измет. Овој вид на пренесување се нарекува „станица на хитгут“ или „стеркорарна“ и е значително помалку ефикасен од преносот на плунковните.
(Одете во Животниот циклус на T. крузи, Екологија на пренос или Стратегии за контрола.)
Песочни мушички и Леишманија

Песочните мушички се вектори за Лајшманија, како и за некои бактериски (Бартонелоза) и вирусни (треска од песочна мушичка) болести. Возрасните песочни мушички имаат должина од околу 2 мм и се карактеризираат со тела и крилја на влакната (слика?). Родови на песок муви падне во две групи: Флеботомус и Сергеантомија најде во Источна хемисфера (Европа, Азија, Африка, Австралија), и Луцомија, Брумптомија и Варилеја во западната хемисфера (т.е. Нов свет). Општо земено, видовите стари светски мушички живеат во пустински или полусуви екосистеми, а видовите на Новиот свет се шумски живеалишта. Некои од старите светски видови ќе се размножуваат во перидоместички ситуации и ќе влезат во човечки живеалишта, додека пренесувањето на болести во Новиот свет е поврзано со луѓе кои живеат или работат во близина на шумата.
Песочните мушички одговорни за преносот на лајшманијазата се Флеботомос и Луцомија, во стариот и во новиот свет. Просторната дистрибуција на лајшманиоза и други пренесени болести на песочни мушички има тенденција да биде лесна заради ограничениот опсег на летање на песокот. Нивното летање често се опишува како „скокање“, се карактеризира со кратки рафали на летови, одделени со неколку секунди одмор. Песочните мушички имаат краток опсег на летови и обично се наоѓаат на десетина метри од местото за размножување.
Како општо правило, само женските песочни мушички земаат оброци во крвта и тие се хранат најактивно на самрак или ноќ, или во услови на слаба осветленост како што е сенка. Тие имаат кратки делови од устата и се хранат за базен. Залак произведува папула во боја на роза опкружена со еритематозна област со дијаметар од околу 10-20 мм.
Макрофагите инфицирани со амастиготи се проголтаат со крвниот оброк и се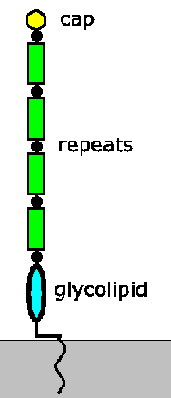 трансформираат во промастиготи (види Циклус на животот на Лајшманија). Овие проциклични промастиготи прикачени на епителот на средното црево на мувата и се реплицираат. Прилогот е со посредство на липофосфогликан (LPG). LPG е изобилство на гликоконјугат на изобилство на клеточна површина составена од три главни домени: сидро на гликолипидна мембрана, повторувања на дисахарид и мало капаче (Слика). LPG-то претрпува биохемиски промени кога паразитите созреваат во метациклични промастиготи. Дисахарид се повторува приближно двојно по број, што доведува до издолжување на LPG и капакот се менува од галактоза во арабиноза. Се верува дека менувањето на структурата на капачето е поврзано со одвојување од епителот на цревата со тоа што се верува дека приврзаноста е поврзана со специфични лектини за галактоза кои се наоѓаат во епителот на цревата на песочната мушичка. Елонготонот на LPG е поврзан со зголемена отпорност на комплементот што укажува на дополнителни улоги за LPG во однос на инфективноста на ‘рбетниот домаќин. (За преглед на гликоконјугатите на Лајшманија, видете: Деско и Турко, Весник за биохемија и биофизика 1455:341; 1999.)
трансформираат во промастиготи (види Циклус на животот на Лајшманија). Овие проциклични промастиготи прикачени на епителот на средното црево на мувата и се реплицираат. Прилогот е со посредство на липофосфогликан (LPG). LPG е изобилство на гликоконјугат на изобилство на клеточна површина составена од три главни домени: сидро на гликолипидна мембрана, повторувања на дисахарид и мало капаче (Слика). LPG-то претрпува биохемиски промени кога паразитите созреваат во метациклични промастиготи. Дисахарид се повторува приближно двојно по број, што доведува до издолжување на LPG и капакот се менува од галактоза во арабиноза. Се верува дека менувањето на структурата на капачето е поврзано со одвојување од епителот на цревата со тоа што се верува дека приврзаноста е поврзана со специфични лектини за галактоза кои се наоѓаат во епителот на цревата на песочната мушичка. Елонготонот на LPG е поврзан со зголемена отпорност на комплементот што укажува на дополнителни улоги за LPG во однос на инфективноста на ‘рбетниот домаќин. (За преглед на гликоконјугатите на Лајшманија, видете: Деско и Турко, Весник за биохемија и биофизика 1455:341; 1999.)
Исто така, постојат фактори во рамките на плунката на песочна мушичка кои ја потенцираат инфективноста на Леишманија кај домаќинот на ‘рбетници. Некои од овие соединенија на плунковните жлезди имаат имуносупресивни активности против лимфоцитите и макрофагите што може да го објаснат потенцирањето.
(Врати се на Леишманија Животот циклус или оди на Контрола стратегии.)
Анофелес и Плазмодиум

Крлежи и Бабезија
Стратегии за контрола
Погледнете специфични активности за контрола на болеста:
ЛИНКИ
- Врвот на страницата
- Табела на вектори
- Це-це и африкански трипанозоми
- Тријатомини и Трипаносома крузи
- Песочни мушички и Леишманија
- Анофелес и плазмодиум
- Крлежи и Бабезија
- Стратегии за контрола
- Протозоологија дом
- Други курсеви и предавања
- Помудар дом
Mark F. Wiser, Tulane University (© 2001). Последно ажурирање на 30 ноември 2020 година.