Original page: http://www.tulane.edu/~wiser/protozoology/notes/INTRO.html
Марк Ф. Уизер (Mark F. Wiser)
Кои се протозои?
Дешифрирање грчки корени резултати во дефинирањето на протозои како “првиот” (прото) “животни” (зоа). Иако молекуларната филогенетска студии покажуваат дека протозои се меѓу првите разгранување еукариотите (види филогенетско дрво), таквата дефиниција не дава многу описни информации. Протозоите не се лесно дефинирани, бидејќи тие се различни и често се оддалечуваат само меѓусебно. Поради екстремната разновидност на протозоа, единствената карактеристика која е заедничка за сите протозои е дека тие се едноклеточни еукариотски микроорганизми. Протозоите поседуваат типични еукариотски органели и воопшто ги покажуваат типичните карактеристики на другите еукариотски клетки. На пример, мембранско врзано јадро кое ги содржи хромозомите се наоѓа кај сите видови протозоа. Меѓутоа, кај многу видови од протозои, некои органели може да бидат отсутни, или пак се морфолошки или функционално различни од оние кои се наоѓаат во други еукариоти. Покрај тоа, многу од протозоите имаат органели кои се единствени за одредена група на протозои.
Прашања кои би можеле да ги постават за дефинирање на протозои вклучуваат: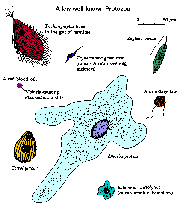
- Што тие изгледа?
- Колку е голема се тие?
- Каде се тие во живо?
- Како тие се движат?
- Како тие се размножуваат?
Протозоите покажуваат широк спектар на морфологии (кликнете на сликата за поголема слика и опис). Не постои ниту една форма или морфологија која би вклучила мнозинство на протозои. Форми се движат од аморфни и постојано менувачки форми на амеба до релативно ригидни форми диктирани делумно од високо наредени цитоскелетони или излачени ѕидови или школки. Неколку протозои видови изразуваат фотосинтетички или други пигменти и на тој начин се обоени. Многу видови протозони покажуваат сложени животни циклуси со повеќе фази. Понекогаш различните фази на животниот циклус се толку различни што ги погрешиле за сосема различни видови.
| Протозојски големини | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
Протозоите – освен неколку колонијални форми – се едноклеточни или едноклеточни организми; иако, некои тврдат дека всушност се “ацелуларни”. Така, огромното мнозинство на протозои се микроскопски. Сепак, тие покажуваат неверојатно голем број на големини. Екстензивните видови се со големина од <1 мкм (10-6 метри) до неколку мм. Се идентификувани фосилизирани форманинферида од неколку см. (Исчезнатите протозои може да се откријат поради излачена калциум карбонатска школка.) Поголемиот дел од организмите што се дискутираат во овој курс ќе бидат 3-50 мкм. Оваа мала големина бара употреба на микроскоп за откривање на протозои. Електронски микроскоп е потребен за детални морфолошки студии.
Протезоите се наоѓаат во влажни средини речиси насекаде. Како група, протозоата се екстремно прилагодливи. Сепак, поединечните видови имаат специфични ниши. Како и сите други организми, протозоите мора да бидат способни да ги стекнат и метаболизираат хранливите материи од нивната средина (т.е. хетеротрофни). Многу протозои едноставно апсорбираат раствори (т.е. осмотрофија) од нивните медиуми, додека некои се чистачи кои внесуваат цврст материјал (т.е. фаготрофија). Грабливните протозои или активно ловат или пасивно ги напаѓаат другите организми (обично бактерии или други протозои). Некои протозои се фотосинтетични и можат да ја освојат енергијата на сонцето и да го претворат во употреблива хемиска енергија (т.е. автоотропни или фотографски). Многу протозои не се ограничени на единствен механизам за хранење и можат да користат комбинации од горенаведените (т.е. миксотрофни).
Протозоите, исто така, може да се гледаат како слободни живи или симбиотички. Општо земено, слободните живи организми се наоѓаат во почвата или во водни средини, додека симбионите живеат во тесна врска со друг организам. Симбиозата подразбира физиолошка зависност на еден организам на друг организам, а не само тесна физичка поврзаност помеѓу два организма. Општо земено оваа зависност е во форма на исхрана. Може да се разликуваат различни форми на симбиоза кои одразуваат природата на асоцијацијата помеѓу двата организма (кутија).
| Симбиотички асоцијации | ||||||
|---|---|---|---|---|---|---|
|
Мотилитетот и цитоскелетонот
| Режими на Мотилитетот | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
Најраните набљудувања на протозои истакна својата подвижност. Ова подвижност резултираше со нивната класификација како “животни”, кои се разликуваат од неподвижни “растенија”. Сепак, подвижност не е универзална карактеристика на протозои и различни протозои користат различни механизми за нивно движење (Табела). Всушност, протозои почетокот биле класифицирани седиште во дел на нивниот механизам на мотилитетот (Види таксономија).
Цилија и флагели се субклелуларни структури кои потикнуваат протозои низ течен медиум. Флагела се долги структури слични на камшици кои го поттикнуваат организмот како резултат на бран-како победи, кој се шири низ нивната должина. Знамените протозои обично имаат една или неколку флагели по организмот. Спротивно на тоа, цилијарните протозои обично се прекриени со редови од бројни цили. Ударите на овие цили се координирани и функционираат како весла за да го поттикнат организмот. Цилија и флагели исто така можат да помогнат во набавката на храна, репродукција и други функции. цилија и флагели се составени од исти компоненти на протеини и се всушност еквивалентни структури. Двете се мембрански врзани филаментозни проекции од ќелијата. Филаментот, познат како аксониме, е составен од серија паралелни микротубули, обично покажувајќи ‘9 + 2’ аранжман. Движењето се создава кога микротубулите се лизгаат едни покрај други. Силата која го посредува ова лизгачко движење е генерирана од протеин наречен динеин. динеин се “моторни протеини” кои ја претвораат хемиската енергија ослободена од АТП хидролиза во механичка енергија. Микротубулите се цитоскелетни елементи кои исто така играат важна улога во клеточната форма и се главна компонента на митотичкото вретено.
За разлика од пливањето изложени од знамиња и цилијати, амеба се протозои кои лазат по цврст супстрат во мода позната како “движење на амебоид”. Амеба проектира псеудоподиум, или лажна нога, од клеточното тело. Псевдопод потоа се прицврстува на супстратот, а потоа го повлекува остатокот од ќелијата на телото напред. Силата вклучена во ова движење е генерирана од друг цитоскелетен систем, кој се состои од актин и миозин. Актин формира долги нишки, исто така познати како микрофиламенти, а миозинот е моторниот протеин кој се движи по микрофиламентите на зависен начин од АТП. Контракцијата на мускулите е уште еден пример за генерирање на сила преку актин-миозински цитоскелетните елементи. Во механистичка смисла, фагоцитозата е форма на амебоидно движење, кое вклучува и микрофиламенти. Во овој случај псевдоподните се прошируваат за да ја опкружуваат честичката која се проголта. Фузија на псеудоподија со телото на клетката резултира со интернализација на честичката во вакуола.
Апикомплекса, исто така, лазат по супстрат, но со различен механизам од амеба. Механизмот на оваа таканаречена “подвижна подвижност” само што почнува да се разбере и веројатно ги вклучува и цитоскелеталните системи базирани на микрофилументи и микротубули. Апикомплекс исто така покажува и интрацелуларни форми, а инвазијата на клеточната клетка исто така ја вклучува и оваа пловидната подвижност. (Види исто така дискусија на клетката на домаќинот инвазија од страна на паразитот на маларијата.)
Мобилната подвижност вклучува генерација на сила преку цитоскелетални елементи базирани на микротубули или цитоскелетни елементи базирани на микрофиламентите. Ова важи и за протозои и други еукариоти. Вклучувањето на микротубулите и микрофиламентите во облик на клеточна форма и движење на клетките ги прават овие субклеточни структури повеќе аналогни на мускулно-скелетниот систем.
Репродукција
| Режими на репродукција | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
Протозоите, како и сите други организми, се репродуцираат. Најчестата форма на репродукција кај протозоите е асексуална бинарна фисија. Со други зборови, еден организам ќе се подели на два еднакви организми. Мала модификација на оваа бинарна фисија, наречена младиот младиот, е кога една од новоформираните клетки е помала од другата. Обично поголемата ќелија се нарекува мајка, а помалата е ќерката. Некои протозои ќе формираат интрацелуларен пупка и во суштина ќе раѓаат. Друга варијација на бинарна фисија е повеќекратна фисија или сегментација. Во оваа ситуација, неколку рунди на нуклеарна репликација се јавуваат без цитокинеза. Оваа мултинуклеарна ќелија тогаш ќе формира повеќе потомства истовремено.
Многу протозои покажуваат сексуална репродукција покрај асексуалните форми на репродукција. Оваа сексуална репродукција може да вклучи производство и фузија на гамети во процеси слични на повисоки организми. Цилиофората е подложена на конјугација во која спротивните типови на парење ќе пар и директно разменуваат генетски материјал (т.е. ДНК). Понекогаш сексуалната репродукција е задолжителен чекор во животниот циклус, додека во други случаи, организмот може да се репродуцира асексуално со повремениот круг на сексуална репродукција.
Во краток преглед, протозои се еукариотски едноклеточни микроорганизми. Сепак, износот на разновидност во поглед на морфологијата, големината и начинот на живеење изложба на протозои го прави тешко да се развие повеќе прецизна дефиниција. Нивниот долг еволутивната историја (види филогенетско дрво) е одговорна за голем дел од оваа разновидност. Сепак, протозои направи изложба карактеристики за сите еукариоти. (Врска до серија на PowerPoint презентации на еукариотска клетка биологија.)
| Потекло на еукариотските клетки |
|---|
| Еволутивниот скок од прокариот во еукариот е значителен и има околу 5000 гени кои се наоѓаат во еукариоти кои не се наоѓаат во прокариот (1). Првите еукариотски организми најверојатно личат на она што сега би го класифицирале како протозои. Меѓутоа, најверојатно нема директен потомци од раните еукариоти и патеката од прокариоти на еукариот најверојатно е оптеретена со неколку недостасува врски. Два клучни настани во еволуцијата на еукариот се веројатно развој на внатрешен цитоскелетален систем и фаготрофија.
Главната разлика помеѓу еукариот и прокариот е крутиот муреин, или пептидогликан, клеточен сид на бактерии. Иако многу еукариоти покажуваат клеточни ѕидови, тие се фундаментално различни од оние на бактериите и веројатно се добиваат секундарно по формирањето на еукариот. На пример, клеточните ѕидови на растителните клетки се состојат главно од целулозни и габични клеточни ѕидови, а ѕидовите на цисти во многу протозои се првенствено составени од хитин. Ригидноста и јачината на клеточните ѕидови ги штитат клетките од механички стрес и даваат осмотска стабилност. Така, совпаѓање со губењето на ѕидот на муреинскиот мобилен сид во раниот еукариот беше развојот на ендоцитоскелетните системи базирани на актин и тубулин за да се компензира релативната кршливост на голи плазма мембрани. Покрај тоа, раните еукариот најверојатно еволуирале во бентосната средина, а особено во густите морските микробиолошки потпирачи. Органските супстанции излачени од бактериите и висока концентрација на сол на таква средина би имале осмопротективен ефект. Густите микробиолошки душеци, исто така, ќе обезбедат средина во која плен е многу изобилен и фагоцитозата може да еволуирала без ефикасна клеточна подвижност. Ескариотот на предците веројатно е поврзан со плен преку клеточни површини со N-врзани гликопротеини кои се развиле по губењето на ѕидот на муреинските клетки, а потоа ги дигерирале однадвор со секретирање на хидролитички ензими. За да се направи процесот поефикасен, примитивните предатори би можеле постепено да го опколат својот плен додека плен целосно не бил зафатен во мембраните на домаќинот со варење што се изведува внатрешно во рамките на фагозомите. Овој ендоцитен механизам исто така ќе бара развој на егзоцитен механизам за одржување на површината на плазматската мембрана. Така, еволуцијата на фагоцитозата не само што вклучуваше проширување на функцијата на актинскиот цитоскелет, туку исто така го возеше развојот на трговијата со мембрани и субклеточните прегради, вклучително и јадрото. Еволуцијата на фаготрофија и ендомембрани не само што отвори нов начин на јадење, туку и овозможи ендосимбиоза и поробување на митохондриите. Раниот еукариот веројатно ја зафатил алфа-протеобактеријата и формираше симбиотична врска која се претвора во асимилација на аеробниот метаболизам на овој организам и развојот на митохондриото. Ова поробување на митохондромот веројатно беше уште еден клучен настан во еволуцијата на еукариотите, бидејќи постоечките еукариоти се добиени од предците што содржат митохондрии. Дополнителните анаеробни еукариоти без митохондрии имаат органели добиени од митохондрии, како што се митозоми и хидрогеноми, или барем имаат нуклеарни гени кои кодираат митохондријални гени. Се претпоставува дека амитохондријалниот еукариотски предок бил во конкурентна неповолна положба и не преживеал долгорочно. Подоцна, цијанобактерија беше поробена од прогениторот на растенијата и се разви во хлоропласти и слично, овие претходници на растенијата беа зафатени и поробени за да ги формираат пластидите пронајдени во многу други алги и протозои. Цитоскелетот продолжил да ги еволуира и ги проширува своите функции, а особено почнал да игра улога во клеточната подвижност. Најзначајно тубулин стана клучна компонента кај флагелата, а подоцна миозинот еволуирал за да овозможи развој на движење на амебоид и движење на пливање. Оваа подвижност им овозможила на овие рани еукариоти да се прошират и во други ниши (на пр., Планктонски, копнени) и затоа овој аеробен, фаготрофски и фланелизиран предци еукариот може да зрачи во разновидната низа протозои и, во крајна линија, другите еукариоти кои во моментов постојат.
|
Таксономија
Таксономија, или систематика, е наука за именување и класификација на организмите. Во прилог на одредување на хиерархиска таксономски класификации, систематика, исто така, се обиде да стави организми во групи одразуваат еволутивен односи или филогени. Сепак, таксономски критериуми често се произволни и таксономија е секогаш се менува за да се рефлектираат новите откритија и толкувања.Исто така, утилитаристички карактеристики, како што се видот на болеста, домаќин опсег и географската дистрибуција, често се користат во систематиката на патогени микроорганизми. Ова е особено точно за протозоа таксономија. Покрај тоа, постои дебата на целокупната филозофија во класификацијата на протозои и односите меѓу многу протозоата групи не се познати (види ДJ Патерсон во Aм. нац. 154, с. 96-124). Класификацијата на протозои е дополнително комплицирана од оригиналната дефиниција на протозои се едноклеточни хетеротропи. Сега се знае дека протозои (или протестира) може да се користат повеќе исхрана стратегии и не може да се смета како едноставно или растителна како (автоотроп) или животно-како (хетеротроп).Така, терминот протозои не може да се смета како вистински таксономска група. И покрај фактот дека зборот протозои, сама по себе не е веќе соодветно таксономско име, тоа е уште една корисна и функционални рок.
Историски протозои беа поделени во четири главни групи: амеба, на флагели, на инфузија и спорозоан. Особините меѓу двете групи беше врз основа на мотилитетот (т.е. амебоид, флагели, влакненцата на клетките). На спорозоан беа хетерогена група, кои произведуваат спори во текот на една фаза на нивниот животен циклус и изложени на “плови” подвижност. Меѓутоа, таквата шема за класификација е сосема произволна и не ги одразуваат вистинската еволутивен односите меѓу организмите.Еден проблем со користење на подвижност како таксономски критериуми е дека многу протозои користат различни типови на подвижност во различни фази на нивниот животен циклус. На пример, неглерия постои во форма на една амеба кога храната е обилен и се претвора во флагелат кога храната е отсутен. Во принцип, на амеба се хетерогена група и сите се веројатно добиени од флагели. Меѓу овие четири оригинални протозоата групи само инфузија се уште се смета за валидна таксон.
Почнувајќи од 1960 година електронски микроскоп беше искористена за да се идентификуваат ултраструктурните карактеристики кои можат да послужат како критериуми за групирање протозои. Во многу случаи морфологија води до класификација што ја става организми во монофилетична групи. Монофилетична значи дека сите на организмите во таа група се веројатно потекнува од еден заеднички предок. На пример, многу од протозои порано наречена спорозоан поседуваат субклеточните структури, колективно познати како апикална органели, и сега се формираат монофилетична група наречена апикомплекс. Сепак, субклеточните структури и метаболички патишта може да се изгуби во некои лози и ставање на тие потомци може да биде проблематично.
Во текот на 1980 година и продолжува до денес молекуларни техники се применуваат на таксономија. Можно еволутивна историја и односи може да се добиени со споредување на ДНК или протеин секвенци. Податоци молекуларна низа потврди филогени врз основа на други критериуми, се населиле некои дебати, и доведе до неколку изненадувања. На пример, молекуларна податоци потврдуваат дека апикомплекс се монофилетична, а освен тоа, укажува на тоа дека тие се поврзани со инфузија и динофлагели. Овие три групи сега се комбинирани во еден поголем монофилетична група наречена алвеолата. Овој однос претходно бил осомничен и за името е во врска со морфолошки структури познати како алвеоларна вреќички. Но, користењето на еден ген може да биде несигурен средство за утврдување еволутивен односи, особено меѓу далечна роднина организми, и молекуларна податоци треба да се толкува со претпазливост.
Од средината на 1980-тите, класификацијата на протестите е во состојба на флукс и се соочува со некои филозофски контроверзии. Од една страна, постојат аргументи за задржување на елементите на бутшлската шема засновани на мотилитет поради неговата блискост и едноставност. Сепак, овие шеми често се во конфликт со филогенетските податоци. Покрај тоа, хиерархиите и редовите на традиционалната систематика Линеа (т.е. филум, класа, со цел, семејство, род, видови) не се вклопуваат добро со микроорганизми и често е тешко да се одлучи кое хиерархиско ниво е најсоодветно за било која посебна протозоја група.
Претходно многу таксономски шеми дефинираа пет царства на животот: прокариоти (бактерии), протести, растенија, габи и животни. Во овие шеми, протозоата се дел од Протиста, заедно со едноклеточни алги, диатоми, оомицети и листести калапи. Сепак, отсекогаш постоело незадоволство од групата протести. Ова делумно се должи на тоа што делумно дефи- ниран критериум се дефинира. Со други зборови, организмите кои не се вклопуваат во другите четири кралства се обврзани во протестот. Покрај тоа, некои протести се филогенетски поблиски поврзани со другите три еукариотски царства отколку со другите протести, а со тоа протестите се јасно полифилетички.
Кавальје-Смит предлага пет еукариотски царства кои се состојат од базал, а со тоа и парафилетички, царство Протозоа и четири изведени кралства: Анималии, Габи, Планта и Хромиста (Слика). Некои од поранешните протести сега се вклучени во изведените царства дека тие се најтесно поврзани со што резултира во царството Протозоа да стане монофилетичар. Сепак, алвеолатите, во кои спаѓаат многу традиционални протозои како што се апикомплекс, цилијати и динофлагелати, формираат клада со хромиста, но сепак се ставаат во протозоанското кралство. Така, хромистите не се холофилетични. Преселувањето на алвеолатите во новото царство наречено хромалвеолата ќе го реши овој проблем. Сепак, ова веројатно ќе резултира со контроверзии и конфузија, бидејќи многу од алвеолатите веќе долго време се сметаат за протозои.
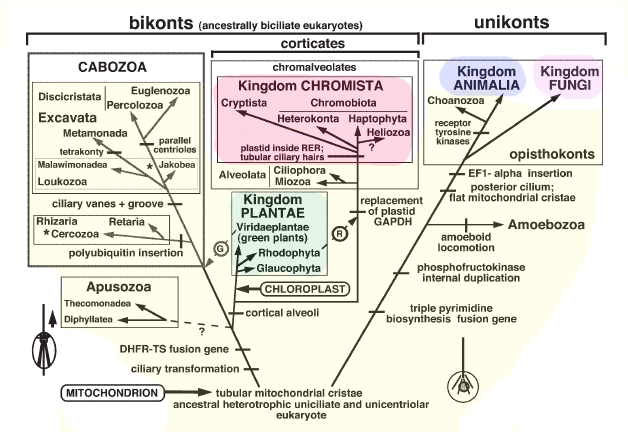
Класификација на еукариот во 5 царства: протозои (жолти), растенија (зелени), хромиста (розова), животни (сини) и габи (лаванда). Модифициран од Кавальје-Смит (2003) Европски журнал за Протистологија 39:338-348.
ЛИНКОВИ
- Кои се протозоа? (врв)
- Таксономија
- Протозоологија насловна
- Еукариотски клеточна биологија
- Други курсеви и предавања
- Помудар насловна
Овие страници се развиени и одржувани од Марк Ф. Уизер, Универзитетот Тулан (© 2000). Последно ажурирање на 25 јули 2017 година.